| |
Instituto de Conservación y Mejora de la Agrodiversidad Valenciana (COMAV) Grupo de Biología Celular |


|
Bases celulares, moleculares y genéticas de la totipotencia celular.Introducción Las células vegetales son totipotentes. Dicho de otro modo, una célula vegetal, más o menos diferenciada, es capaz de revertir su proceso de diferenciación y volver a un estado primigenio, desdiferenciado, de tipo meristemático, sin especialización concreta pero capaz de especializarse en cualquier otro tipo celular. Esta fascinante capacidad se manifiesta en la naturaleza en forma de diversas estructuras que la planta forma como respuesta a heridas, ataques, o simplemente como mecanismo de reproducción asexual. Por ejemplo, cuando un esqueje accidentalmente arrancado enraíza espontáneamente en el suelo, o cuando brotan nuevos individuos a partir de bulbos, tubérculos o rizomas. Estas estructuras naturales vienen siendo utilizadas por los agricultores para propagar vegetativamente las plantas de sus cultivos cuando la reproducción por semillas presenta dificultades, o no es tan rentable, en términos económicos, como la asexual. Pero además, esta propiedad abre vías experimentales muy interesantes para poder explotarla con fines biotecnológicos. Si sabemos cómo revertir el proceso natural de diferenciación, y posteriormente inducir específicamente la diferenciación hacia el tipo celular que nosotros deseemos, estaremos en disposición de conseguir la regeneración de plantas completas, siempre que las condiciones sean las adecuadas. Si conocemos estas condiciones, es decir, los mecanismos tanto celulares como moleculares o genéticos que gobiernan esta capacidad de las células vegetales de revertir a un estado indiferenciado, y después entrar en un nuevo proceso morfogénico para dar lugar a nuevos tipos celulares, órganos e incluso individuos, seremos capaces de poder controlar estos procesos, e inducirlos a voluntad cuando queramos, en aras de mejorar los cultivos ya existentes, o de producir nuevas variedades más adaptadas al mercado, a la aparición de nuevas plagas y enfermedades, a las nuevas condiciones derivadas del cambio climático, o a cualquier otra circunstancia que ahora no pudiera preverse. Ejemplos de la utilidad de este tipo de estudios en el ámbito de la agricultura y la mejora vegetal hay muchos y variados. Muchos de ellos tienen que ver con la reproducción asexual típica de las plantas. Además de aprovechar las estructuras naturalmente generadas para la reproducción asexual, antes mencionadas, el ser humano ha aprendido a inducir de forma artificial la multiplicación asexual para propagar vegetativamente plantas a su antojo. Para ello se vale desde hace siglos de distintas técnicas agronómicas basadas en la totipotencia y la capacidad regenerativa asexual de las plantas como el estaquillado, el acodado o la división de rizomas, bulbos o tubérculos. Sin embargo, el abanico de posibilidades biotecnológicas que se ha abierto gracias a los cultivos in vitro de tejidos o células aisladas ha revolucionado la forma en que actualmente se entiende la producción vegetal, al permitir la multiplicación masiva de material vegetal en poco espacio, menos tiempo y con una notable reducción de los costes de producción. Además de este aspecto, muy relevante, el cultivo in vitro tiene muchas más aplicaciones como método de reproducción asexual. De hecho, el cultivo in vitro podría definirse como el conocimiento de este tipo de recetas para manipular la totipotencia de las células vegetales. Esta poderosa herramienta permite la propagación de grandes volúmenes de plantas en espacios reducidos y en mucho menor tiempo que con técnicas tradicionales de cultivo. Por otro lado, es de gran utilidad en la obtención de plantas libres de patógenos, plantas 100% homozigotas, en la producción de plantas en peligro de extinción, en estudios de ingeniería genética, fitorremediación, etc. De hecho, de acuerdo con los trabajos publicados en las revistas más influyentes del campo de la biología vegetal, se podrían establecer cinco áreas principales en las que los cultivos in vitro están siendo aplicados:
Más concretamente, dentro del contexto de la agricultura, horticultura y silvicultura, se pueden identificar siete grandes áreas de utilidad del cultivo in vitro:
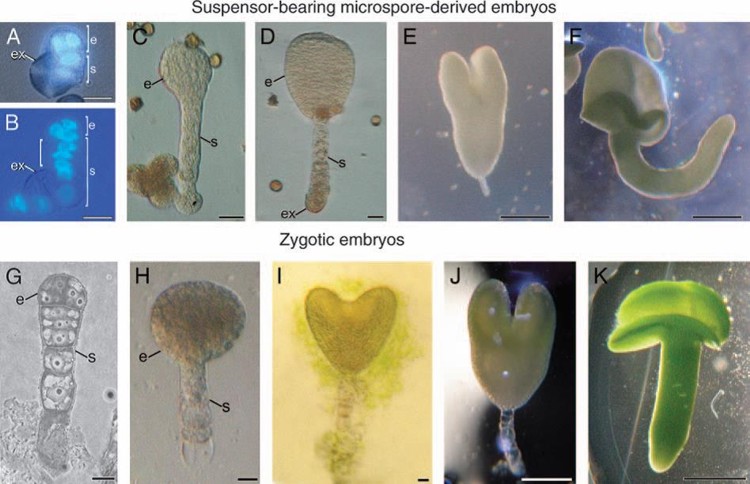
De todas estas áreas, la mejora vegetal es la disciplina donde el cultivo in vitro ha aportado y está aportando más, siendo hoy en día una herramienta insustituible. A partir de la década de los 70, los métodos in vitro se han venido utilizando frecuentemente como un complemento a los métodos tradicionales de mejora vegetal. En la actualidad, su uso ya no es complementario, sino imprescindible para muchas facetas de la mejora. Por ejemplo, para generar variabilidad de origen somaclonal para la obtención, por ejemplo, de flores, hojas o frutos de colores o formas nuevas, producción o desaparición de pigmentos, variegaciones, tamaños o portes nuevos o infrecuentes, distintas lobulaciones en las hojas, etc. Más allá de la variación somaclonal, existen muchas otras aplicaciones del cultivo in vitro en la mejora vegetal que implican explantes de naturaleza tanto somática (no implicados ni directa ni directamente en la reproducción sexual) como germinal. Dentro de los explantes de naturaleza somática, podemos mencionar la embriogénesis somática, la manipulación de la ploidía para obtener, entre otros, tetraploides, o la fusión de protoplastos. La embriogénesis somática consiste en la inducción a partir de células somáticas del desarrollo de un embrión completo, y perfectamente capacitado para germinar y dar lugar a una nueva planta, genéticamente idéntica a la donante del explante. Es decir, está técnica abre la puerta a la obtención de semillas artificiales, como ya está ocurriendo en diversas especies. Los tetraploides son individuos con el genoma duplicado que tienen distintas ventajas en cuanto a tamaño, producción y otros caracteres de interés agronómico. La fusión de protoplastos proporciona a la mejora vegetal una valiosa herramienta para abordar la superación de barreras a la hibridación interespecífica de una forma radicalmente distinta a cualquier otro abordaje. Dentro de los explantes de naturaleza germinal, quizá uno de los mejores ejemplos sería la obtención de individuos haploides y dobles haploides. Un individuo haploide es aquel que proviene de una célula inicialmente haploide. Por definición, las únicas células haploides de un organismo son los gametos, o sus precursores, una vez pasada la meiosis. Por tanto, los individuos haploides provendrán de los gametos, o de las microsporas o megasporas que los originan. De forma intencionada o no, estos individuos pueden sufrir una duplicación de su genoma haploide. Sea inducida o no inducida, lo que se obtiene tras la duplicación del genoma es un individuo doble haploide, cuyo genoma está formado por dos copias exactas del genoma inicialmente haploide. Es decir, en cada uno de los cromosomas homólogos habrá exactamente los mismos alelos. Serán homocigotas para todos y cada uno de sus loci. En términos genéticos, esto implica que los individuos doble haploides tendrán siempre la misma descendencia de autofecundación, no generarán jamás poblaciones segregantes para ningún carácter. Por más recombinación que sufran los gametos de los dobles haploides, siempre se generarán las mismas combinaciones de genes, al haber los mismos alelos en cada cromosoma homólogo. Esto, claro está, será así en ausencia de mutaciones o de cualquier otro fenómeno generador de variabilidad más allá de la recombinación. En términos de mejora genética, a estos individuos genéticamente homogéneos se les denomina líneas puras. Es decir, un doble haploide es una línea pura, y esta es precisamente su gran ventaja, pues es lo que necesitan las empresas de producción de semillas para generar sus híbridos comerciales, pero obtenido de forma mucho más rápida y económica. Esta alternativa reduce las típicas 7-8 generaciones de autofecundación necesarias para estabilizar un genotipo híbrido a tan solo una in vitro. Resulta pues, mucho más rápida y económica. En mejora vegetal, estas líneas son también imprescindibles para la cartografía genética de caracteres complejos como producción o calidad, que son los de mayor interés agronómico y no son abordables hoy día mediante otras técnicas. Constituyen también una poderosa herramienta en transgénesis, para evitar hemizigotos y ahorrar tiempo y recursos en la obtención de plantas transformadas con el transgen en ambos alelos. Por otra parte, desde un punto de vista científico básico, estas líneas son también muy útiles para estudios básicos de ligamiento y estimas de fracciones de recombinación. Aunque estos estudios pueden también realizarse convencionalmente (retrocruces o F2), las líneas dobles haploides tienen la ventaja de ser autoperpetuables, es decir se pueden perpetuar simplemente a partir de semillas de autofecundación. Son también una herramienta sumamente útil para la selección genética y la detección de mutantes recesivos, pues el fenotipo de las plantas resultantes no se ve afectado por los efectos de la dominancia, y los caracteres determinados por genes recesivos pueden ser fácilmente identificados. Otra ventaja es la posibilidad de servir como sistema modelo para el estudio del desarrollo embriogénico in vitro, sin la interferencia del tejido materno, pues la embriogénesis cigótica y la haploide derivada de microsporas presentan un gran número de semejanzas (Figura 1).
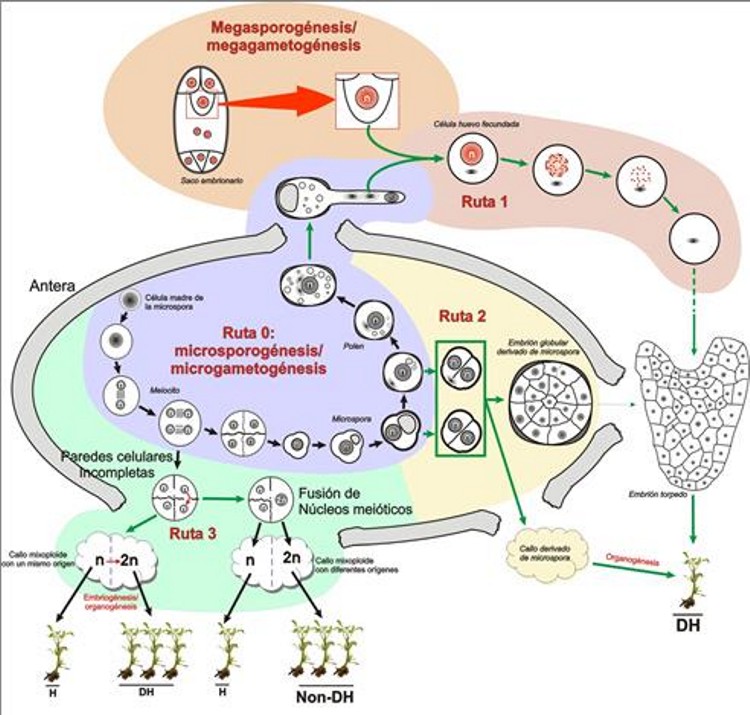
Es posible obtener haploides y doble haploides a través de diferentes vías de desarrollo, tanto del gametófito masculino como del femenino. A partir del gametófito femenino es posible obtener haploides y doble haploides través de una vía conocida como ginogénesis. También es posible mediante determinados tipos de hibridaciones interespecíficas entre especies cercanas, o utilizando polen al que se le han aplicado tratamientos especiales, como la irradiación. Pero la más interesante, por su eficiencia y por el número de especies en que es aplicable, es la posibilidad de obtener haploides y doble haploides a partir del gametófito masculino, a través de la inducción de una serie de rutas experimentales que se engloban dentro de un fenómeno conocido como la androgénesis. Androgénesis La androgénesis se define como el proceso de generación de un individuo cuyo fondo genético proviene exclusivamente de un núcleo de origen masculino (Seguí-Simarro, 2010). Es decir, consiste en la generación de una planta a partir exclusivamente de un gameto masculino o de su precursor haploide (gametofito). Existen distintas vías para inducir la androgénesis. Tal como se ilustra en la Figura 2, puede formarse un individuo haploide androgénico in vivo mediante la fecundación de la célula huevo y posterior inactivación del genoma del núcleo femenino (ruta 1 de la Figura 2), o in vitro a través de organogénesis sobre callos derivados de meiocitos (ruta 3 de la Figura 2), o induciendo la embriogénesis (o en algunos casos callogénesis) a partir de microsporas (ruta 2 de la Figura 2).
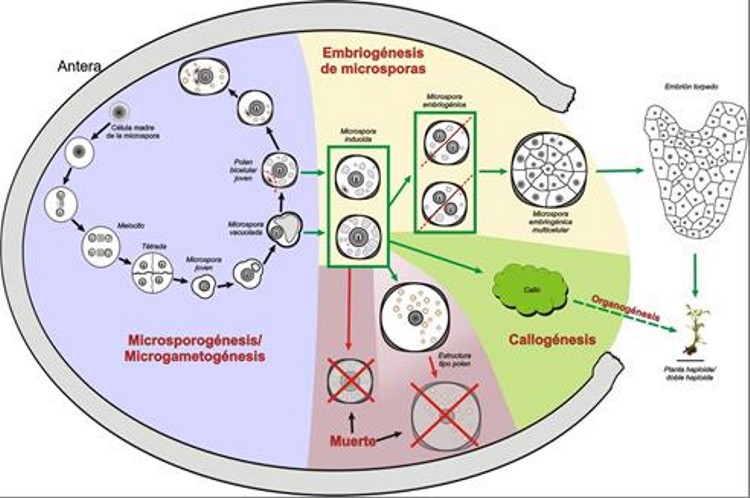
De entre ellas, la más útil y por ello ampliamente utilizada en la gran mayoría de especies donde se pueden obtener dobles haploides es la embriogénesis de microsporas, es decir, la reprogramación hacia embriogénesis de las microsporas (inicialmente destinadas a ser granos de polen), dando lugar a embriones dobles haploides, que tienen un enorme interés práctico para la producción de líneas puras con las que obtener variedades híbridas comerciales. Esta ruta experimental, alternativa al desarrollo normal del polen, fue descubierta por Guha y Maheshwari (1964). Utilizando experimentalmente esta ruta de desarrollo es posible obtener líneas puras, dobles haploides, mediante la inducción de embriogénesis o de callogénesis haploide partiendo generalmente del gametófito masculino inmaduro (la microspora). Las microsporas se pueden desviar de la ruta gametofítica e inducir a formar un embrión o un callo haploide de dos maneras (Seguí-Simarro y Nuez, 2008a): o bien aislándolas de la antera y cultivándolas en medio de cultivo líquido (cultivo de microsporas), o bien cultivando directamente la antera que las contiene en medio sólido (cultivo de anteras). En cualquiera de los dos casos, tendremos un sistema en el que coexisten distintas poblaciones de microsporas (Figura 3). Habrá una población de microsporas que son desviadas de su desarrollo natural y son efectivamente inducidas a proliferar, convirtiéndose en embriones y posteriormente germinando para dar lugar a una planta completa. En algunas especies, como berenjena, los embriones se transforman en callos, pero pueden también obtenerse plantas de ellos. El porcentaje de microsporas que adoptan esta vía varía mucho según la especie, pero en general, es por desgracia minoritario. Otro porcentaje de ellas no se inducirá a proliferar, y expresará un programa de desarrollo similar al del polen, dando lugar a estructuras de tipo polen, que finalmente, acabarán muriendo. Y finalmente, encontraremos una mayoría de microsporas que o bien llegaron ya muertas al cultivo in vitro, o no fueron capaces de resistir el estrés inherente al tratamiento de inducción y antes o después acabarán muriendo. En definitiva, habrá una mayoría de microsporas o muertas o no embriogénicas junto con una minoría sí inducida.
La embriogénesis se puede inducir en microsporas de diversas especies de angiospermas, tanto mono como dicotiledóneas, y gracias a ella se pueden regenerar plantas completas a través de embriogénesis haploide directa o indirectamente a través de una fase intermedia de callo, mediante técnicas comunes de cultivo in vitro. Se obtendrán, por tanto, plantas haploides en las que se induce posteriormente la duplicación cromosómica (Seguí-Simarro y Nuez, 2008b), o directamente dobles haploides (por duplicación no inducida del genoma haploide). En ambos casos, plantas 100% homocigotas. Sin embargo, esta ruta de desarrollo no puede inducirse con igual eficiencia en todas las especies y variedades. Hay algunas que son extremadamente recalcitrantes, y otras que son muy fácilmente inducibles, como la colza. Para el estudio y el avance en el conocimiento del fenómeno de la embriogénesis de microsporas, la colza (Brassica napus) ha tenido un papel fundamental. La colza es una crucífera de enorme importancia a nivel mundial en la producción de aceite a partir de sus semillas oleaginosas. Pero además, por su elevada respuesta a los tratamientos inductores, algunas líneas de B. napus son sistemas modelo para el estudio de la inducción y desarrollo de los embriones androgénicos (Chupeau et al., 1998; Friedt y Zarhloul, 2005; Seguí-Simarro y Nuez, 2008a). Este sistema se ha demostrado a lo largo de los años como el más eficiente desde un punto de vista aplicado y fácil desde un punto de vista experimental. De hecho, es en esta especie donde se han conducido la mayoría de estudios sobre la inducción de la embriogénesis de microsporas y el desarrollo del embrión haploide, generando resultados aplicables a la mejora del proceso en otras especies menos sensibles. Dado que en este sistema es donde más datos se tienen a nivel celular y molecular, resulta más fácil y útil la profundización en su estudio. Esta aproximación viene siendo utilizada con éxito por nuestro grupo desde su inicio. De hecho, poseemos una amplia experiencia en el uso de este sistema modelo para el estudio de este fenómeno. En los últimos años, hemos estado estudiando, entre otros aspectos, el papel que juega la autofagia (Corral-Martínez et al., 2013; Parra-Vega et al., 2015a) y el estrés de retículo endoplásmico en la inducción de embriogénesis, y más recientemente, el papel de la pared celular de las microsporas embriogénicas en la resiliencia al estrés (Parra-Vega et al., 2015b; Corral-Martínez et al., 2019; Rivas-Sendra et al., 2019), y el papel del calcio intracelular en la señalización y/o activación de la inducción de este proceso de forma diferente en especies/líneas de alta y baja respuesta (Rivas-Sendra et al., 2017; Rivas-Sendra et al., 2019). Objetivo de la línea de investigación Todas las posibilidades biotecnológicas basadas en la totipotencia celular que hemos descrito en el apartado anterior, y en particular la obtención de dobles haploides, tienen una enorme importancia aplicada al sector agrobiotecnológico, como también se ha mencionado. Sin embargo, es todavía poco lo que se sabe de muchas de ellas. De hecho, y volviendo al ejemplo concreto de la androgénesis, aunque sería aplicable a muchos otros, hay todavía especies donde sería muy interesante disponer ya de estas tecnologías puestas a punto, pero donde por desgracia aún no se ha avanzado lo suficiente para que esto sea una realidad transferible al sector industrial. Para avanzar en dicha mejora, en los casi 50 años de investigación en DHs se han utilizado dos tipos de abordajes experimentales. La vía más fácil y directa, aunque quizá no la más eficiente, es la utilizada hasta ahora en la mayoría de los laboratorios que tratan de optimizar este tipo de protocolos en especies o variedades concretas: probar distintas sustancias, sospechosas de tener un efecto positivo, y evaluar si realmente tienen dicho efecto. Nuestra experiencia durante años utilizando este enfoque nos indica que, aunque se han conseguido avances, esta estrategia quizá no sea la más eficiente, pues solo algunas sustancias han demostrado tener un papel relevante, y otras han sido descartadas. Así pues, consideramos que es necesario dar un giro a la manera en que tradicionalmente se viene abordando este problema, y que es fundamental potenciar aproximaciones basadas en el conocimiento. Es decir, se hace necesario profundizar en los mecanismos biológicos subyacentes a este fenómeno. Por ello, una de las principales líneas de investigación de nuestro grupo es el estudio de los distintos factores implicados en el proceso de inducción. Utilizando un abordaje experimental paralelo basado en la combinación de técnicas de biología celular, molecular y genética, tratamos de detectar marcadores de inducción útiles para identificar aquellas microsporas efectivamente inducidas, y tratamos de identificar nuevos factores implicados en la inducción, aplicables a mejorar la inducción en otras especies, más recalcitrantes. Bibliografía Corral-Martínez P., Driouich A. y Seguí-Simarro J.M. (2019). Dynamic Changes in Arabinogalactan-Protein, Pectin, Xyloglucan and Xylan Composition of the Cell Wall During Microspore Embryogenesis in Brassica napus. Front Plant Sci 10. Corral-Martínez P., Parra-Vega V. y Seguí-Simarro J.M. (2013). Novel features of Brassica napus embryogenic microspores revealed by high pressure freezing and freeze substitution: evidence for massive autophagy and excretion-based cytoplasmic cleaning. J. Exp. Bot. 64, 3061-3075. Chupeau Y., Caboche M. y Henry Y. (1998). Androgenesis and haploid plants. (Berlin, Heidelberg.: Springer-Verlag). Friedt W. y Zarhloul M.K.(2005). Haploids in the improvement of crucifers. En Haploids in crop improvement II, C.E. Palmer, W.A. Keller y K.J. Kasha, eds (Berlin: Springer-Verlag), pp. 191-213. Guha S. y Maheshwari S.C. (1964). In vitro production of embryos from anthers of Datura. Nature 204, 497. Parra-Vega V., Corral-Martínez P., Rivas-Sendra A. y Seguí-Simarro J.M.(2015a). Formation and excretion of autophagic plastids (plastolysomes) in Brassica napus embryogenic microspores. Front Plant Sci 6 (94). Parra-Vega V., Corral-Martínez P., Rivas-Sendra A. y Seguí-Simarro J.M. (2015b). Induction of embryogenesis in Brassica napus microspores produces a callosic subintinal layer and abnormal cell walls with altered levels of callose and cellulose. Front Plant Sci 6, 1018. Rivas-Sendra A., Calabuig-Serna A. y Seguí-Simarro J.M. (2017). Dynamics of Calcium during In vitro Microspore Embryogenesis and In vivo Microspore Development in Brassica napus and Solanum melongena. Front Plant Sci 8, 1177. Rivas-Sendra A., Corral-Martínez P., Porcel R., Camacho-Fernández C., Calabuig-Serna A. y Seguí-Simarro J.M. (2019). Embryogenic competence of microspores is associated with their ability to form a callosic, osmoprotective subintinal layer. J. Exp. Bot. 70, 1267–1281. Seguí-Simarro J.M. (2010). Androgenesis revisited. Bot. Rev. 76, 377-404. Seguí-Simarro J.M. y Nuez F. (2008a). How microspores transform into haploid embryos: changes associated with embryogenesis induction and microspore-derived embryogenesis. Physiol. Plant. 134, 1-12. Seguí-Simarro J.M. y Nuez F. (2008b). Pathways to doubled haploidy: chromosome doubling during androgenesis. Cytogenet. Genome Res. 120, 358-369. |
|
© 2019 Grupo de Biología Celular. |
Aviso Legal |