| |
Instituto de Conservación y Mejora de la Agrodiversidad Valenciana (COMAV) Cell Biology Group |


|
Cellular, molecular and genetic bases of cell totipotency.Introduction Plant cells are totipotent. In other words, a more or less differentiated plant cell is able to revert its differentiation process and return to a previous, dedifferentiated, meristematic-like state, without defined specialization but capable of specializing in any other cell type. This fascinating ability is manifested in nature in the form of different structures that plants form in response to wounds, attacks, or simply as a mechanism of asexual reproduction. For example, when an accidentally detached cutting roots spontaneously on the ground, or when new individuals sprout from bulbs, tubers or rhizomes. These natural structures are used by farmers to vegetatively propagate the plants of their crops when seed reproduction presents difficulties, or is not as profitable, in economic terms, as asexual reproduction. In addition, this property opens up very interesting experimental pathways to be exploited for biotechnological purposes. If we know how to revert the natural process of differentiation, and subsequently how to specifically induce differentiation towards the cell type we want, we will be able to achieve the regeneration of whole plants, provided that the conditions are appropriate. If we know these conditions, that is, the cellular, molecular or genetic mechanisms that govern this ability of plant cells to revert to an undifferentiated state, and then enter a new morphogenic process to give rise to new cell types, organs and even individuals, we will be able to control these processes, and induce them at will whenever we want, in order to improve existing crops, or to produce new varieties more adapted to the market, to the emergence of new pests and diseases, to the new conditions derived of climate change, or any other circumstance that can not be currently foreseen. Examples of the usefulness of this type of studies in the field of agriculture and plant improvement are many and varied. Many of them have to do with the typical asexual reproduction of plants. In addition to taking advantage of the naturally generated structures for asexual reproduction, mentioned above, the human being has learned to artificially induce asexual multiplication to vegetatively propagate plants at will. For this, humans have been taking advantage for centuries of different agronomic techniques, based on totipotency and the asexual regenerative capacity of plants, such as stabbing, bending or division of rhizomes, bulbs or tubers. However, the range of biotechnological possibilities that plant cell and tissue in vitro culture has opened is revolutionizing the way in which plant production is currently understood, by allowing massive multiplication of plant material in a reduced space, less time and with a notable reduction in production costs. In addition to this very relevant aspect, in vitro culture has many more applications as a method for asexual reproduction. In fact, in vitro culture could be defined as the knowledge of this type of recipes to manipulate the totipotence of plant cells. This powerful tool allows for the propagation of large volumes of plants in confined spaces and in much less time than with traditional cultivation techniques. On the other hand, it is very useful in obtaining pathogen-free plants, 100% homozygous plants, in the production of endangered plants, in genetic engineering studies, phytoremediation, etc. In fact, according to the work published in the most influential journals in the field of plant biology, five main areas in which in vitro cultures are being applied could be established:
More specifically, within the context of agriculture, horticulture and forestry, seven large areas of usefulness of in vitro culture can be identified:
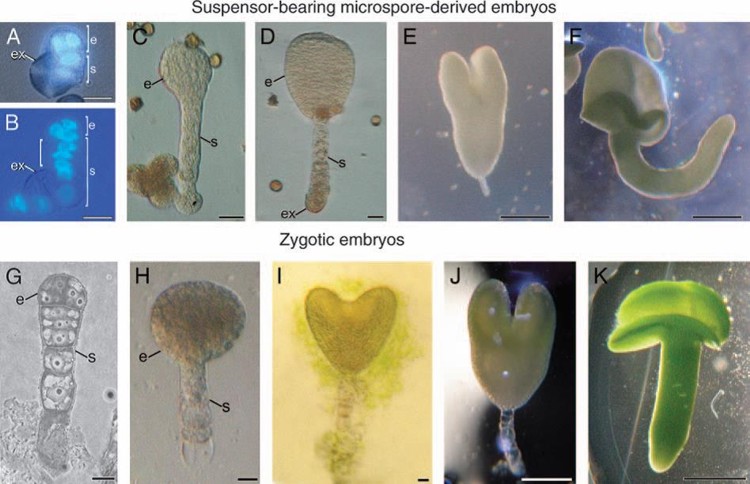
Among all these areas, plant breeding is the discipline where in vitro culture has contributed and is contributing more, being today an irreplaceable tool. Since the 1970s, in vitro methods have been frequently used as a complement to traditional plant breeding methods. At present, its use is no longer complementary, but essential for many facets of improvement. For example, to generate variability of somaclonal origin to obtain, for example, flowers, leaves or fruits of new colors or shapes, production or elimination of pigments, variegations, new or infrequent sizes or shapes, different lobulations on the leaves, etc. Beyond somaclonal variation, there are many other applications of in vitro culture in plant breeding that involve explants of both somatic nature (not directly or directly involved in sexual reproduction) and germinal. Within the explants of somatic nature, we can mention somatic embryogenesis, the manipulation of ploidy to obtain, among others, tetraploids, or the fusion of protoplasts. Somatic embryogenesis consists in the induction from somatic cells of the development of a complete embryo, perfectly capable of germinating and giving rise to a new plant, genetically identical to the donor of the explant. That is, this technique opens the door to obtaining artificial seeds, as is already being done in various species. Tetraploids are individuals with the doubled genome that have different advantages in terms of size, production and other traits of agronomic interest. Protoplast fusion provides plant breeding with a valuable tool to address the overcoming of barriers to interspecific hybridization in a way radically different from any other approach. Within the germinal explants, perhaps one of the best examples would be to obtain haploid and doubled haploid individuals. A haploid individual is one that comes from a haploid cell. By definition, the only haploid cells in an organism are gametes, or their precursors, having passed meiosis. Therefore, haploid individuals will come from gametes, or from the microspores or megaspores that originate them. Intentionally or not, these individuals may undergo a duplication of their haploid genome. Whether induced or not induced, what is obtained after genome duplication is a doubled haploid individual, whose genome is made up of two exact copies of the initially haploid genome. That is, there will be exactly the same alleles in each of the homologous chromosomes. They will be homozygous for each and every one of their loci. In genetic terms, this implies that doubled haploid individuals will always have the same self-fertilization offspring, they will never generate segregating populations for any character. Along generations and recombination rounds, the same combinations of genes will always be generated in doubled haploid gametes, as there are the same alleles in each homologous chromosome. This, of course, will be so in the absence of mutations or any other phenomenon that generates variability beyond recombination. In terms of genetic improvement, these genetically homogeneous individuals are called pure lines. That is, a doubled haploid is a pure line, and this is just its great advantage, as it is what seed production companies need to generate their commercial hybrids, but obtained much faster and more economically. This alternative reduces the typical 7-8 generations of self-fertilization needed to stabilize a hybrid genotype to just one in vitro. It is therefore much faster and cheaper. In plant breeding, these lines are also essential for the genetic mapping of complex characters such as production or quality, which are those of greatest agronomic interest and are not approachable today by other techniques. They are also a powerful tool in transgenesis, to avoid hemizygotes and save time and resources in obtaining plants transformed with the transgene in both alleles. On the other hand, from a basic scientific point of view, these lines are also very useful for basic studies of linkage and estimates of recombination fractions. Although these studies can also be carried out conventionally (backcross or F2), doubled haploid lines have the advantage of being self-perpetuating, that is, they can simply be perpetuated from self-fertilization seeds. They are also an extremely useful tool for the genetic selection and detection of recessive mutants, since the phenotype of the resulting plants is not affected by the effects of dominance, and the characters determined by recessive genes can be easily identified. Another advantage is the possibility of serving as a model system for the study of embryogenic development in vitro, without the interference of maternal tissue, since zygotic embryogenesis and haploid microspore embryogenesis have a large number of similarities. (Figure 1).
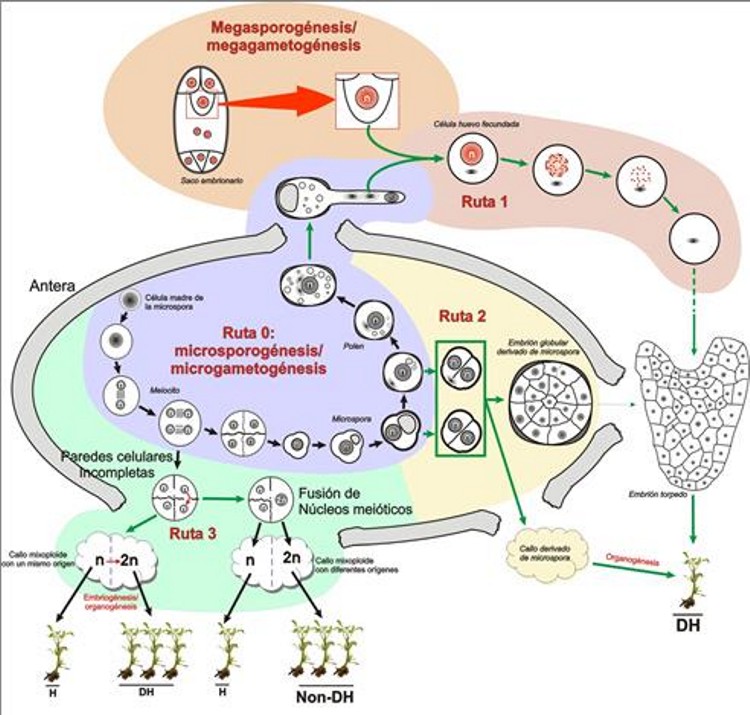
It is possible to obtain haploids and doubled haploids through different developmental pathways, from both the male and the female gametophyte. From the female gametophyte it is possible to obtain haploids and doubled haploids through a pathway known as gynogenesis. It is also possible through certain types of interspecific hybridizations between phylogenetically close species, or using pollen irradiated with ionizing radiations. But the most interesting, due to its efficiency and the number of species in which it is applicable, is the possibility of obtaining haploids and doubled haploids from the male gametophyte, through the induction of a series of experimental routes that are included within a phenomenon known as androgenesis. Androgenesis Androgenesis is defined as the process of generating an individual whose genetic background comes exclusively from a nucleus of male origin (Seguí-Simarro, 2010). That is, it consists in the generation of a plant exclusively from a male gamete or its haploid precursor (gametophyte). There are different ways to induce androgenesis. As illustrated in Figure 2, an androgenic haploid individual can be formed in vivo by fertilization of the egg cell and subsequent inactivation of the female nucleus genome (route 1 of Figure 1), or in vitro through organogenesis on meiocyte-derived calli (route 3 of Figure 2), or inducing embryogenesis (or in some cases callogenesis) from microspores (route 2 of Figure 2).
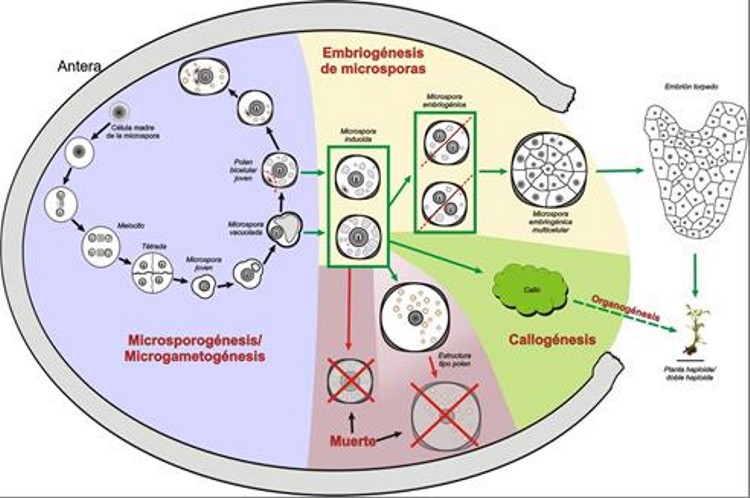
Among them, the most useful and therefore widely used in the vast majority of species where doubled haploids can be obtained is microspore embryogenesis, that is, reprogramming towards embryogenesis of microspores (initially intended to be pollen grains), giving rise to doubled haploid embryos, which have an enormous practical interest for the production of pure lines with which to obtain commercial hybrid varieties. This experimental route, an alternative to normal pollen development, was first discovered by Guha and Maheshwari (1964). Using this experimental developmental route, it is possible to obtain pure, doubled haploid lines, by inducing haploid embryogenesis or callogenesis, generally starting from the male gametophyte precursor (the microspore). Microspores can be deviated from the gametophytic pathway and induced to form an embryo or haploid callus by two techniques (Seguí-Simarro and Nuez, 2008a): either by isolating them from the anther and growing them in liquid culture medium (microspore culture), or by directly cultivating the anther that contains them in solid medium (anther culture). In either case, we will have a system where different microspore populations coexist (Figure 3). There will be a population of microspores that are diverted from their natural development and are effectively induced to proliferate, becoming embryos and subsequently germinating to give rise to a complete plant. In some species, such as eggplant, embryos become calli, but plants can also be obtained from them. The percentage of microspores that adopt this route varies greatly depending on the species, but unfortunately, they are minority. Another percentage of them will not be induced to proliferate, and will express a developmental program similar to that of pollen, resulting in pollen-like structures, which will eventually die. Finally, we will find a majority of microspores that were either already dead upon in vitro culture initiation, or not able to withstand the stress inherent to induction treatments, and sooner or later they will end up dying. In short, there will be a majority of microspores either dead or non-embryogenic, together with a minority of induced microspores.
Embryogenesis can be induced in microspores of various angiosperm species, both monocots and dicots, and whole plants can be regenerated through haploid embryogenesis directly or indirectly through an intermediate phase of callus by common in vitro culture techniques. Either way gives rise to haploid plants in which chromosomal duplication is subsequently induced (Seguí-Simarro and Nuez, 2008b), or directly doubled haploid plants (by non-induced duplication of the haploid genome). In both cases, 100% homozygous plants will be produced. However, this developmental pathway cannot be induced with the same efficiency in all species and varieties. There are some that are extremely recalcitrant, and others that are very easily inducible, such as rapeseed. For the study and advancement in the knowledge of the phenomenon of microspore embryogenesis, rapeseed (Brassica napus) has played a fundamental role. Rapeseed is a cruciferous of enormous importance worldwide in the production of oil from its oilseeds. But in addition, due to their high response to inductive treatments, some B. napus lines constitute models systems for the study of the induction and development of androgenic embryos (Chupeau et al., 1998; Friedt and Zarhloul, 2005; Seguí-Simarro and Nuez, 2008a). This system has proven over the years as the most efficient from an applied point of view and easy from an experimental point of view. In fact, it is in this species where most studies on induction of microspore embryogenesis and development of the haploid embryo have been conducted, generating results applicable to the improvement of the process in other, less sensitive species. Since this system is where more data are available at the cellular and molecular level, it is easier and more useful to deepen its study. This approach has been used successfully by our group since its inception. In fact, we have extensive experience in using this model system to study this phenomenon. In recent years, we have been studying, among other aspects, the role played by autophagy (Corral-Martínez et al., 2013; Parra-Vega et al., 2015a) and endoplasmic reticulum stress in embryogenesis induction, and more recently, the role of the cell wall of the embryogenic microspore in stress resilience (Parra-Vega et al., 2015b; Corral-Martínez et al., 2019; Rivas-Sendra et al., 2019), and the role of intracellular calcium in signaling and/or activation of this process differently in species/lines with high and low responses (Rivas-Sendra et al., 2017; Rivas-Sendra et al., 2019). Objective of the research line All the biotechnological possibilities based on cellular totipotency described in the previous section, and in particular doubled haploid technology, have an enormous importance applied to the agrobiotechnological sector, as also mentioned. However, little is known about many of them. In fact, and going back to the particular example of androgenesis, although it would be applicable to many others, there are still species where it would be very interesting to have these technologies already developed, but where, unfortunately, not enough progress has yet been made for this to be a reality transferable to the industrial sector. To gain knowledge in this topic, two types of experimental approaches have been used in the almost 50 years of research in DHs. The easiest and most direct approach, although perhaps not the most efficient, is the one used so far in most laboratories that try to optimize this type of protocol in specific species or varieties: to test different substances, suspected of having a positive effect, and to evaluate if they really have such an effect. Our experience for years using this approach indicates that, although progress has been made, this strategy may not be the most efficient, since only some substances have proven to have a relevant role, and others have been discarded. Thus, we consider that it is necessary to turn the way in which this problem has been traditionally addressed, and that it is essential to enhance knowledge-based approaches. That is, it is necessary to deepen the biological mechanisms underlying this phenomenon. Therefore, one of the main lines of research of our group is the study of the different factors involved in the induction process. Using a parallel experimental approach based on the combination of cell biology, molecular biology and genetics, we try to detect induction markers useful to identify those effectively induced microspores, and try to identify new factors involved in induction, applicable to improve induction in other, more recalcitrant species. References Corral-Martínez P., Driouich A. and Seguí-Simarro J.M. (2019). Dynamic Changes in Arabinogalactan-Protein, Pectin, Xyloglucan and Xylan Composition of the Cell Wall During Microspore Embryogenesis in Brassica napus. Front Plant Sci 10. Corral-Martínez P., Parra-Vega V. and Seguí-Simarro J.M. (2013). Novel features of Brassica napus embryogenic microspores revealed by high pressure freezing and freeze substitution: evidence for massive autophagy and excretion-based cytoplasmic cleaning. J. Exp. Bot. 64, 3061-3075. Chupeau Y., Caboche M. and Henry Y. (1998). Androgenesis and haploid plants. (Berlin, Heidelberg.: Springer-Verlag). Friedt W. and Zarhloul M.K.(2005). Haploids in the improvement of crucifers. En Haploids in crop improvement II, C.E. Palmer, W.A. Keller and K.J. Kasha, eds (Berlin: Springer-Verlag), pp. 191-213. Guha S. and Maheshwari S.C. (1964). In vitro production of embryos from anthers of Datura. Nature 204, 497. Parra-Vega V., Corral-Martínez P., Rivas-Sendra A. and Seguí-Simarro J.M.(2015a). Formation and excretion of autophagic plastids (plastolysomes) in Brassica napus embryogenic microspores. Front Plant Sci 6 (94). Parra-Vega V., Corral-Martínez P., Rivas-Sendra A. and Seguí-Simarro J.M. (2015b). Induction of embryogenesis in Brassica napus microspores produces a callosic subintinal layer and abnormal cell walls with altered levels of callose and cellulose. Front Plant Sci 6, 1018. Rivas-Sendra A., Calabuig-Serna A. and Seguí-Simarro J.M. (2017). Dynamics of Calcium during In vitro Microspore Embryogenesis and In vivo Microspore Development in Brassica napus and Solanum melongena. Front Plant Sci 8, 1177. Rivas-Sendra A., Corral-Martínez P., Porcel R., Camacho-Fernández C., Calabuig-Serna A. and Seguí-Simarro J.M. (2019). Embryogenic competence of microspores is associated with their ability to form a callosic, osmoprotective subintinal layer. J. Exp. Bot. 70, 1267–1281. Seguí-Simarro J.M. (2010). Androgenesis revisited. Bot. Rev. 76, 377-404. Seguí-Simarro J.M. and Nuez F. (2008a). How microspores transform into haploid embryos: changes associated with embryogenesis induction and microspore-derived embryogenesis. Physiol. Plant. 134, 1-12. Seguí-Simarro J.M. and Nuez F. (2008b). Pathways to doubled haploidy: chromosome doubling during androgenesis. Cytogenet. Genome Res. 120, 358-369. |
|
© 2019 Cell Biology Group. |
Legal Notice |