| |
Instituto de Conservación y Mejora de la Agrodiversidad Valenciana (COMAV) Cell Biology Group |


|
Applications of Cell Biology in Plant Biotechnology.
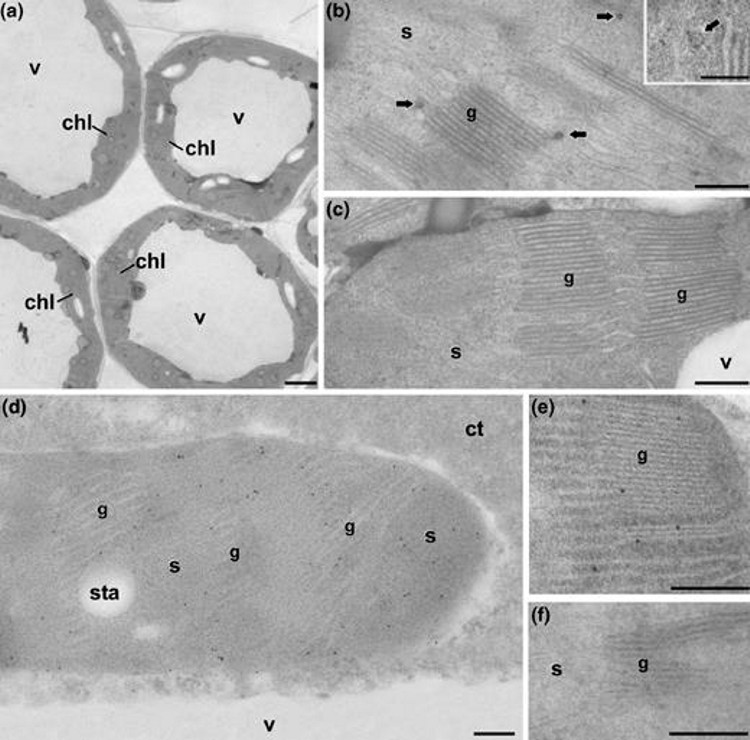
Another research line of our group involves the application of the cell biology techniques available in our Laboratory, to gain knowledge or to solve specific problems related to plant biology and biotechnology. In particular, our group has years of experience in the application of techniques such as in vitro culture, flow cytometry, light and fluorescence microscopy, transmission and scanning electron microscopy to the study of plant material. We are also experts in analytic techniques based on microscopy, such as immunolocalization (immunohistochemistry and immunocytochemistry), fluorescence (FISH) and ultrastructural (electron microscopy) or in situ hybridization. We also handle image analysis, quantification and three-dimensional reconstruction and modeling techniques based on microscopy. In addition to our own lines, in our laboratory we have established a number of scientific collaborations with research groups within and outside Spain, centered on the use of these techniques for different topics of interest. Some of them are illustrated next. Following this philosophy, we are open to any proposal for collaboration with other groups, beneficial in scientific terms to all participants. Identification of therapeutic biomolecules in biofactory plants Between 2007 and 2011 we collaborated with various groups in the identification of various proteins of therapeutic interest in transgenic plant tissues and cells, using molecular in situ localization techniques. These proteins are produced by genetically modified plants, engineered for the production of these biomolecules. Our job includes their in situ detection in those tissues, cells or organelles responsible for their production. For example, in 2008 one of these works in collaboration with the Institute of Agrobiotechnology of the Public University of Navarra (Fernandez-San Millan et al., 2008) was published in a leading journal in the field of Plant Biotechnology (Plant Biotechnology Journal). This study demonstrated the immunogenicity of L1 proteins produced in tobacco plants. The L1 protein is a component of the capsid of human papilloma virus, responsible for women’s cervix cancer. In this work, transgenic tobacco plants expressing large amounts of L1 in their chloroplasts were obtained. Our collaboration was the identification, using specific antibodies, of this protein in the chloroplasts of transgenic plants (Figure 1).
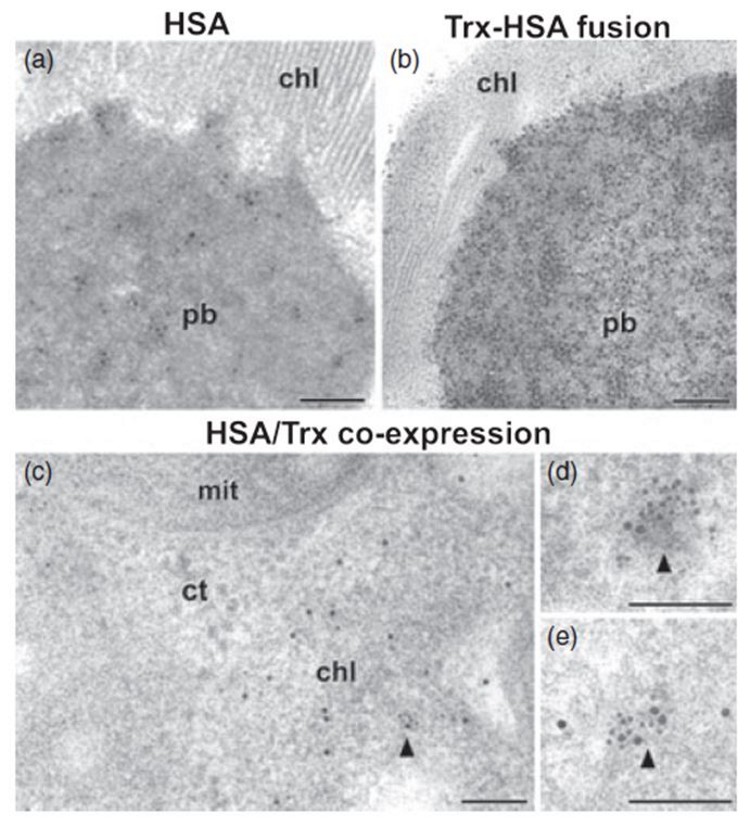
A similar methodological approach allowed us to identify other recombinant proteins of therapeutic interest such as human serum albumin or endogenous thioredoxins in transgenic tobacco chloroplasts (Figure 2). The results have been published in Plant Biotechnology Journal (Sanz-Barrio et al., 2013).
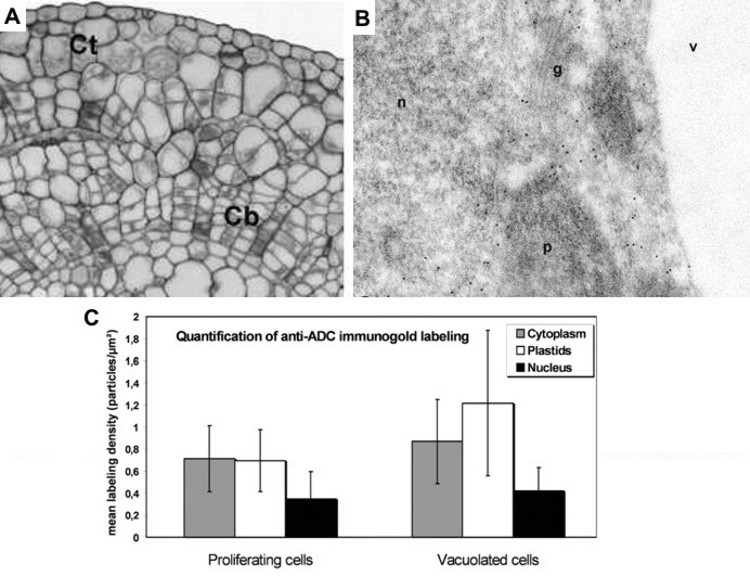
In situ localization of arginine decarboxylase (ADC) during organogenic nodule formation in hop (Humulus lupulus L.) In this collaboration with researchers from the Plant Systems Biology Lab, Center for Biodiversity, Functional & Integrative Genomics (BioFIG), ICAT, University of Lisbon in Portugal, we identified and quantified the presence and abundance of the ADC enzyme in hop organogenic nodules at different stages of development, once induced in vitro (Figure 3). The results have been published in Plant Signaling & Behavior (Fortes et al., 2011).
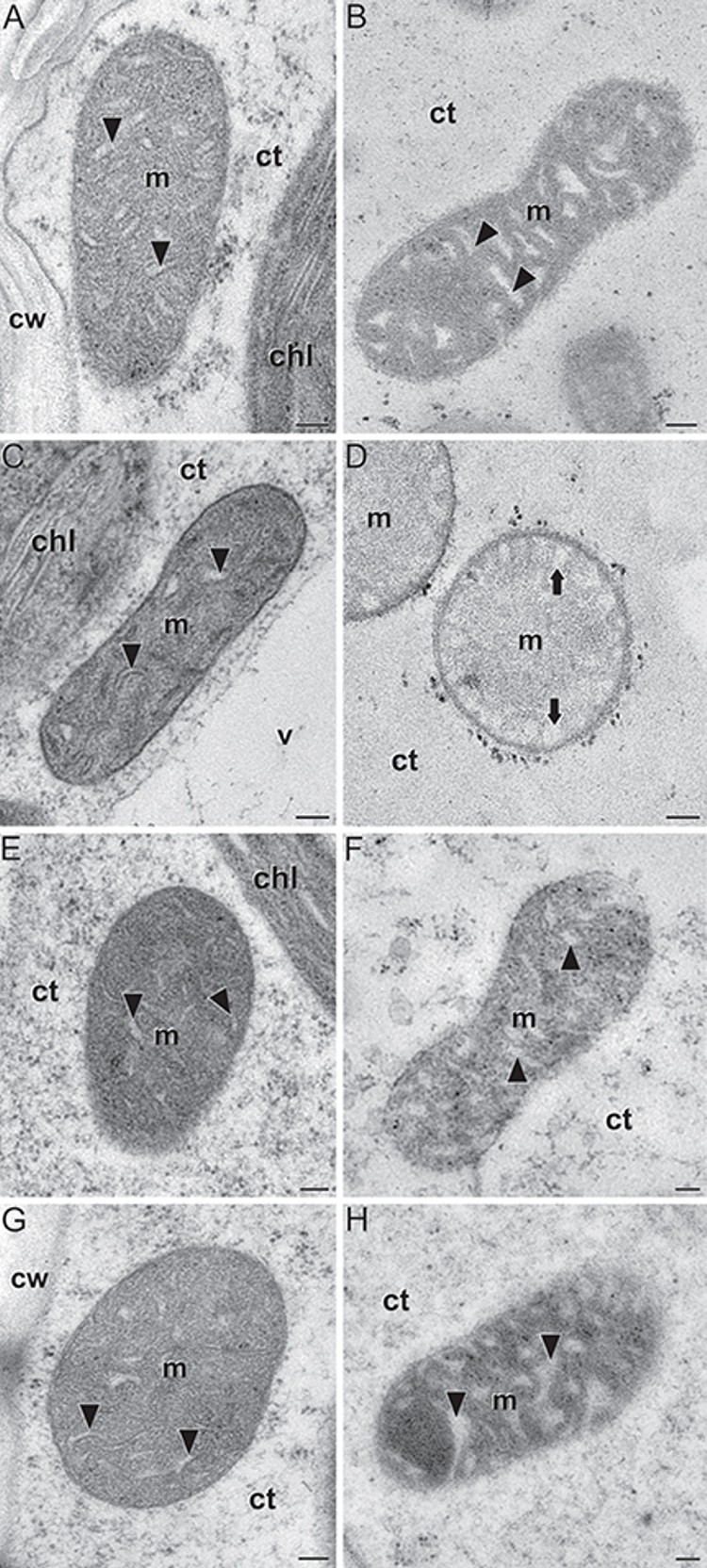
In situ localization of Zea mays Brittle1-1 (ZmBT1-1) in endosperms of wild type and zmbt1-1 mutants in maize (Zea mays) In this collaboration with researchers from the group of Prof. Javier Pozueta at Universidad Pública de Navarra, Spain, we characterized the ultrastructure of mitochondria from endosperms of wild type individuals and different zmbt1-1 maize mutants (Figure 4). The ZmBT1-1 enzyme is an essential component of the starch biosynthetic machinery in maize endosperms. Although traditionally it was thought that its location was plastidial, this work shows that ZmBT1-1 is also located in mitochondria, and that its absence or alteration in the zmbt1-1 mutants induce, among other features, important morphological and ultrastructural alterations, mainly at the level of mitochondrial cristae. The results have been published in Frontiers in Plant Science (Bahaji et al., 2019).
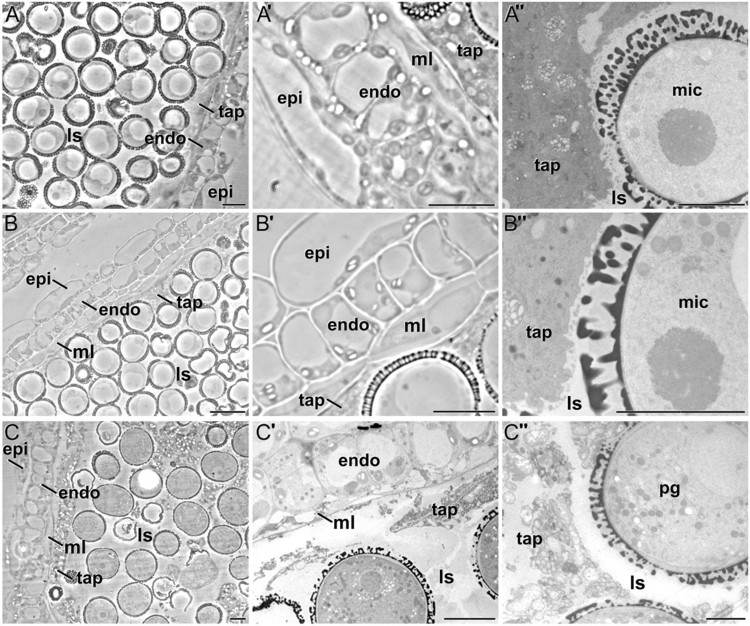
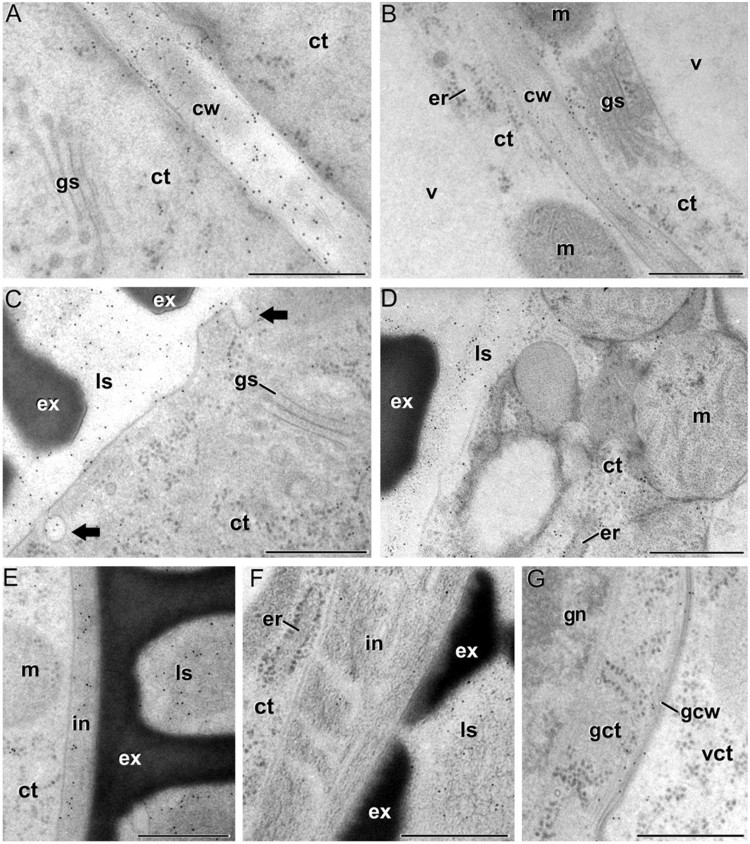
Changes in the levels and distribution of different components of the cell wall of microspores, pollen and the different anther tissues during anther development in Brassica napus In this work, carried out in collaboration with the group of Prof. Aseddine Driouich fron Université de Rouen (France), we performed an extensive and detailed analysis of the changes in cell wall composition during Brassica napus anther development (Figures 5 and 6). We used immunogold labeling to study the spatial and temporal patterns of the composition and distribution of different arabinogalactan protein (AGP), pectin, xyloglucan and xylan epitopes in high-pressure-frozen/freeze-substituted anthers, quantifying and comparing their relative levels in the different anther tissues and developmental stages. The results have been published in Plant and Cell Physiology (Corral-Martínez et al., 2016).
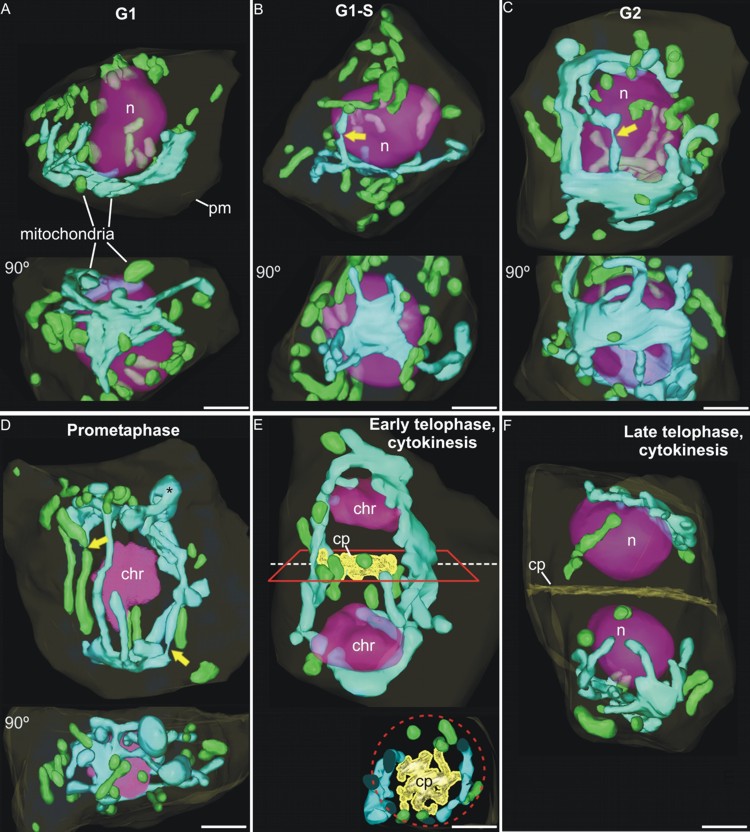
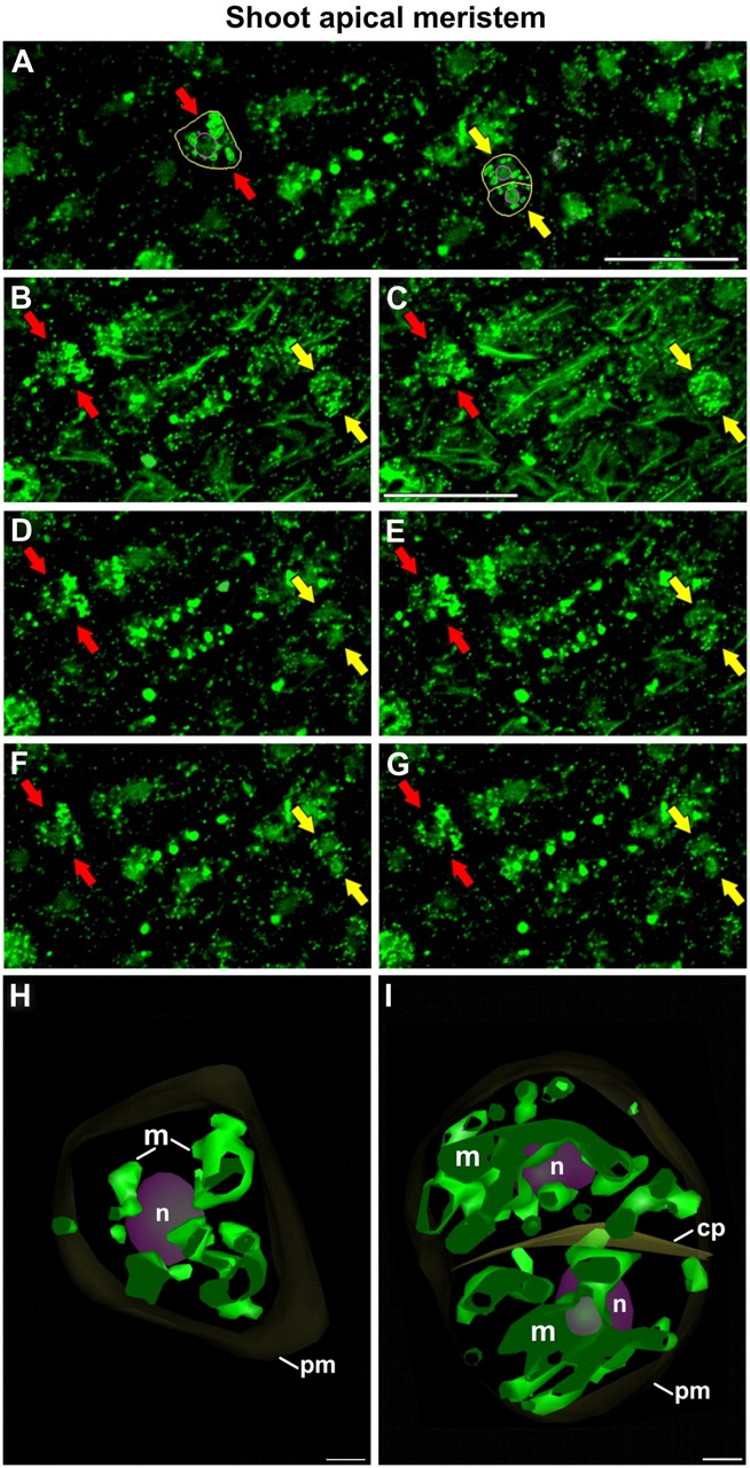
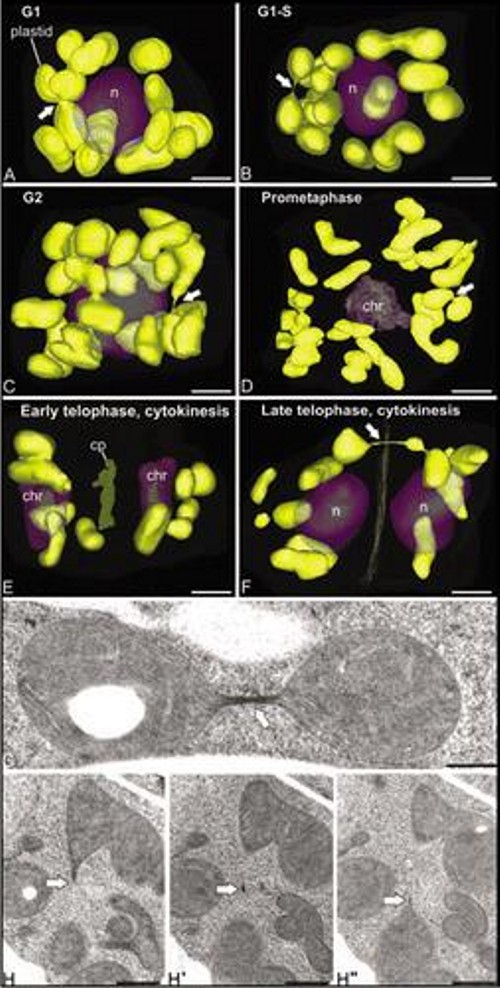
Three-dimensional study of the morphological and distributional changes of the mitochondrial network of Arabidopsis thaliana shoot apical meristem cells In this work, in collaboration with Prof. Andrew Staehelin, from University of Colorado (Boudler, Colroado, USA), we studied the presence of a special type of reticulated mitochondria, forming a dynamic three-dimensional network (Figure 7) around the nucleus of certain meristematic cell types, characterized by their high rate of proliferation and undifferentiated state. For this, we used three-dimensional reconstruction techniques from ultra-thin serial sections observed by transmission electron microscopy (Figure 7), and from confocal slices obtained by confocal laser scanning microscopy (Figure 8). The presence of these mitochondria, never before described in higher plants, and their associated mitochondrial cycle, has important implications for both cellular energetics and plant reproduction, since reticulated mitochondria form a common space within which the mitochondrial DNA (mtDNA) can be homogenized and recombine, so that daughter cells receive a mtDNA copy as homogeneous as the original. However, other DNA-containing organelles such as chloroplasts did not show any morphology or distribution different from the conventional and widely described (Figure 9). These works were published in Plant Physiology (Seguí-Simarro et al., 2008) and Plant Signaling and Behavior (Seguí-Simarro and Staehelin, 2009).
References Bahaji A., Muñoz F.J., Seguí-Simarro J.M., Camacho-Fernández C., Rivas-Sendra A., Parra-Vega V., Ovecka M., Li J., Sánchez-López Á.M., Almagro G., Baroja-Fernández E. and Pozueta-Romero J. (2019). Mitochondrial Zea mays Brittle1-1 Is a Major Determinant of the Metabolic Fate of Incoming Sucrose and Mitochondrial Function in Developing Maize Endosperms. Front Plant Sci 10. Corral-Martínez P., García-Fortea E., Bernard S., Driouich A. and Seguí-Simarro J.M. (2016). Ultrastructural immunolocalization of arabinogalactan protein, pectin and hemicellulose epitopes through anther development in Brassica napus. Plant Cell Physiol. 57, 2161-2174. Fernandez-San Millan A., Ortigosa S.M., Hervas-Stubbs S., Corral-Martinez P., Seguí-Simarro J.M., Gaetan J., Coursaget P. and Veramendi J. (2008). Human papillomavirus L1 protein expressed in tobacco chloroplasts self-assembles into virus-like particles that are highly immunogenic. Plant Biotechnol. J. 6, 427-441. Fortes A.M., Costa J., Santos F., Seguí-Simarro J.M., Cordeiro A., Palme K., Altabella T., Tiburcio A.F. and Pais M.S. (2011). Arginine Decarboxylase expression, polyamines biosynthesis and reactive oxygen species during organogenic nodule formation in hop. Plant Signaling and Behavior In press. Sanz-Barrio R., Corral-Martinez P., Ancin M., Segui-Simarro J.M. and Farran I. (2013). Overexpression of plastidial thioredoxin f leads to enhanced starch accumulation in tobacco leaves. Plant Biotechnol. J. 11, 618-627. Seguí-Simarro J.M., Coronado M.J. and Staehelin L.A. (2008). The mitochondrial cycle of Arabidopsis shoot apical meristem and leaf primordium meristematic cells is defined by a perinuclear tentaculate/cage-like mitochondrion. Plant Physiol. 148, 1380-1393. Seguí-Simarro J.M. and Staehelin L.A. (2009). Mitochondrial reticulation in shoot apical meristem cells of Arabidopsis provides a mechanism for homogenization of mtDNA prior to gamete formation. Plant Signaling and Behavior 4, 168-171. |
|
© 2019 Cell Biology Group. |
Legal Notice |