| |
Instituto de Conservación y Mejora de la Agrodiversidad Valenciana (COMAV) Cell Biology Group |


|
Doubled haploid in recalcitrant solanaceae of agronomic interest.Doubled haploids At present there are protocols available for doubled haploid production through induction of androgenesis in several hundred species of agricultural interest, from grasses such as wheat, barley, rice, rapeseed, tobacco or maize (reviewed in Maluszynski et al., 2003) to woody species as mandarin, bitter orange or oak, among others (reviewed in Srivastava and Chaturvedi, 2008). However, except in model species such as rapeseed, tobacco or barley, the efficiency of androgenic doubled haploid production is still very low (Palmer and Keller, 2005; Seguí-Simarro and Nuez, 2008a; Touraev et al., 2001). This is even more critical in vegetables of high agronomic interest, as those of the Solanaceae family. Of the five major solanaceous crops (pepper, tobacco, potato, eggplant and tomato), only in tobacco sufficient progress have been made to consider this species as a model. Indeed, methods to efficiently produce tobacco double haploids by both anther and isolated microspore cultures are already available, working with an acceptable efficiency (Chupeau et al., 1998; Maluszynski et al., 2003; Touraev et al., 2001). In potato, protocols to induce microspore cultures are also available (Říhová and Tupy, 1999). However, the rest of Solanaceae are considered recalcitrant, as although in some cases doubled haploids have been obtained, we are still far from the efficiency observed in tobacco or rapeseed. In eggplant and pepper only anther culture seems to work, at least for some cultivars, and in tomato none of the two methods is available up to now. Despite the genetic proximity of these five solanaceous species, they seem to respond very differently to androgenic induction. In particular, three of them, tomato (Solanum lycopersicum), eggplant (Solanum melongena) and pepper (Capsicum annuum) are especially recalcitrant. Thus, it is necessary to deepen the study of these systems for a better understanding of the process, to find new ways of induction of androgenesis, to improve the efficiency of the methods in practical application, and if none of these approaches work, to use other, alternative approaches not based on induction of androgenesis. This is the case of tomato, where approaches based on in vitro culture seem not to work efficiently, and CENH3-based alternatives for uniparental chromosome elimination seem more promising. In view of our results in recent years, we can conclude that obtaining doubled haploids from recalcitrant Solanaceae such as tomato, eggplant and pepper, is possible but not easy. This will be our main challenge in this research line. Doubled haploids in tomato Tomato is the first vegetable crop in terms of production and cultivated area, both globally and in the European Union. In Spain, tomato is the main vegetable. However, and despite of its importance, little progress in the tomato DH field as been reported, with no reliable and standardized methods available so far. Over the past 30 years, most of the published research reported the induction of calli or multicellular structures, or the regeneration of roots or apical shoots. Only two laboratories, including ours, have published the complete regeneration of full plants with a haploid or DH origin (Shtereva et al., 1998; Zagorska et al., 1998; Seguí-Simarro and Nuez, 2005, 2006, 2007; Corral-Martínez et al., 2011). In contrast to the high morphological variability of regenerants published previously, our regenerants have high morphological homogeneity, but mixoploidy in a percentage of them and a low general efficiency is still observed (Seguí-Simarro and Nuez, 2007). It is obvious that tomato is extremely recalcitrant, and a much more in-depth study is still essential to obtain successful results. Between 2005 and 2011, our group contributed significantly to the understanding of the problem of androgenesis induction in this crop (Seguí-Simarro and Nuez, 2005, 2006, 2007, Corral-Martínez et al. 2011). Until then, the pollen developmental stage in which this process can be induced in tomato was not clear. We demonstrated that it can be induced in two different stages, although with greatly different results and implications. On the one hand, it is possible to induce meiocytes (Figure 1), before the end of tetrad walling, as long as they are in vitro cultured within the anthers (Seguí-Simarro and Nuez, 2005; Corral-Martínez et al., 2011). This way, haploid, DH, and mixoploid calli can be induced from immature meiocytes (Seguí-Simarro and Nuez, 2007). Moreover, the need for being cultured together with their anther makes possible the occasional appearance of somatic regenerants, coming from the anther wall tissues, not 100% homozygous and therefore not desirable for our purposes. These phenomena, inherent to this technology, represent a major problem when it comes to translating this experimental methodology to production technology, because it requires the genetic evaluation of each single regenerant to determine their origin and ploidy.
Moreover, and as a result of the same investigations, we have also shown that when microspores at the vacuolate stage are isolated and grown in liquid medium, it is possible to induce divisions embryogenic in these microspores (Seguí-Simarro and Nuez, 2007). This alternative would avoid the problems mentioned above, since it eliminates the influence of somatic tissue from the anther and embryos are obtained directly. Microspore-derived embryos are much more genetically stable and less prone to chromosomal instability that microspore-derived calluses (Seguí-Simarro and Nuez 2008b). However, up to now very few genotypes that have been evaluated, and in the few positive results obtained, it has not been possible to go beyond the first divisions in the early development of microspore-derived embryos. For reasons we currently ignore, these embryos arrest, and never progress beyond the globular stage (Seguí-Simarro and Nuez, 2007). Our work in tomato was pioneer in Spain in this field and shed light over some of the most important questions that existed regarding androgenesis in tomato. On the one hand, the meiocyte was confirmed as an inducible stage, but its limited practical application was demonstrated as well. On the other hand, this work opened a way to the production of DHs through the induction of isolated microspores, which we demonstrated as possible, and the subsequent development of embryos. However, this topic is still largely at its infancy, and much more efforts must be devoted. In parallel, the pioneering works of Ravi and Chan in Arabidopsis (Ravi and Chan, 2010) showed that it is possible to obtain haploids and therefore doubled haploids using a completely different approach to those based on in vitro culture and induction of androgenesis or gynogenesis, used until then. In 2010, these researchers demonstrated that through manipulation of the centromeric histone H3 gene (CENH3) it was possible to produce individuals with altered versions of this protein which, when crossed with a wild type (non transformed) individual, produced a diploid zygote with instability in the chromosomes contributed by the mutant parental, which eventually led to the elimination of these altered chromosomes and ended up giving rise to a haploid individual only with the genome of the wild type parental. This new approach to haploidy (and subsequently to production of doubled haploids by chromosome doubling) has opened the door to its study and practical application in other species such as barley (Sanei et al., 2011) or maize (Kelliher et al., 2016). This, together with the development of gene editing tools through CRISPR/Cas technology, is allowing the development of these types of approaches with a greater degree of precision. This is one of the research lines we are currently working with in tomato. Doubled haploids in eggplant Eggplant is another of the most important vegetables worldwide. In Europe, Spain is the leading exporter and second largest producer. This species appears to respond better to the induction of embryogenesis, and DH lines of some varieties and hybrids have been developed (Rotino, 1996; Rivas-Sendra et al., 2017). However, at present only anther cultures seem to work routinely. The results of both ours and many other groups have shown that eggplant embryos can be induced from microspores cultured within the anther. Indeed, our group has been producing doubled haploids for years through anther cultures in different eggplant commercial hybrids and different materials at different stages of breeding programs (Figure 2).
However, anther culture does not exclude the occasional appearance of somatic embryos from anther tissues, as also mentioned for tomato. In addition, we must take into account the uncontrollable secretory effect of the tapetal layer that surrounds the pollen sac, and that prevents us from having a strict control of the culture conditions. In addition, it is well known that anther cultures have a very low efficiency, producing only a few embryos per anther cultivated. In those species where isolated microspore cultures are well set up, it is possible to get hundreds or even thousands of embryos from the microspores contained in a single anther. As in tomato, the development of a method for androgenesis induction from isolated eggplant microspores would be highly desirable, and would avoid the problems mentioned above of tapetum effect, appearance of somatic regenerants, and low efficiency. In our group we have made significant progresses in this topic (Corral-Martínez and Seguí-Simarro, 2012, 2014; Rivas-Sendra et al., 2015), developing a protocol by which we are achieving isolated microspore induction and doubled haploid regeneration (Figure 3) with higher efficiency, well above that previously published.
In 2017 we obtained the first eggplant line highly responsive to microspore culture (Rivas-Sendra et al., 2017) and currently, we try to optimize the protocol even more. In parallel, we try to solve one of the problems that still limit the routine application of this methodology. In the earliest stages of microspore proliferation, they develop to form structures similar to globular embryos, but instead of experiencing the transition from radial symmetry to bilateral symmetry and the consequent differentiation of the different organs typical of zygotic embryos, androgenic embryo cells continue to proliferate in an undifferentiated manner, forming callus-like structures. Although it is possible to obtain haploid and doubled haploid plants by inducing organogenic regeneration in these calli to obtain complete plants, there is no doubt that this process would be faster and easier if we managed to get the embryos to complete their entire development beyond the globular stage, so that they would be ready to germinate autonomously. This is another aspect in which we work at present. Doubled haploids in pepper Pepper is the third solanaceous crop that could be defined as recalcitrant to the induction of androgenesis. Spain uses to rank first or among the first countries of the European Union both in production and cultivated area of pepper, which gives an idea of the importance that an efficient method to obtain doubled haploid lines would have in economic terms. In pepper, the situation is similar to eggplant. For years, anther cultures have been used as a tool for basic studies (Seguí-Simarro et al., 2006; Parra-Vega et al., 2013a; Parra-Vega et al., 2013b) and for applied pepper breeding programs. In fact, as in eggplant, our group has been collaborating and currently collaborates with companies and research institutions in the production of doubled haploids through anther cultures in different eggplant commercial hybrids and different materials at different stages of breeding programs (Figure 4).
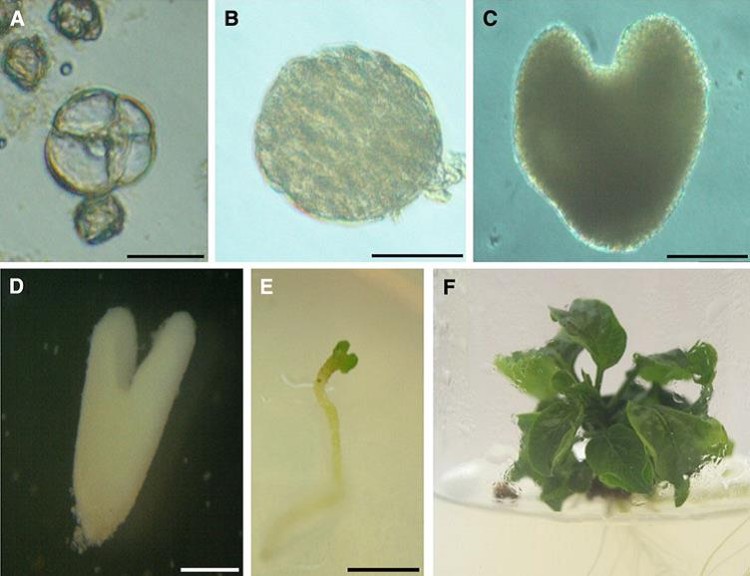
Worldwide, several hybrid seed companies use this technique to obtain pure lines. Obviously, this technique also involves the problems described in the case of eggplant (tapetum effect, the possibility of somatic regenerants and low efficiency), so it is also desirable to achieve a protocol for obtaining DH through isolated microspore culture. In this respect, in the literature there are several approaches to microspore culture, such as the direct isolation (Supena et al., 2006b, Kim et al., 2008) or the shed-microspore approach, which consists of culturing the anthers in a biphasic (solid-liquid) medium, so as to promote anther dehiscence and microspore shedding (Supena et al. 2006a; Supena et al., 2006b). This way, microspores would be isolated from the anther and trapped in the solid-liquid interface. Some of these approaches have shown a limited efficiency (Supena et al., 2006b). Others have proven efficient for embryo production, but not for the quality of these embryos (Kim et al., 2008). The experience of our group is that it is possible to induce the proliferation of pepper microspores to embryos using a method similar to that of eggplant (Figure 5), although more effort is necessary for this methodology to become applicable on a routine basis.
Corral-Martínez P., Nuez F. and Seguí-Simarro J.M. (2011). Genetic, quantitative and microscopic evidence for fusion of haploid nuclei and growth of somatic calli in cultured ms1035 tomato anthers. Euphytica DOI: 10.1007/s10681-010-0303-z. Corral-Martínez P. and Seguí-Simarro J.M. (2012). Efficient production of callus-derived doubled haploids through isolated microspore culture in eggplant (Solanum melongena L.). Euphytica 187, 47-61. Corral-Martínez P. and Seguí-Simarro J.M. (2014). Refining the method for eggplant microspore culture: effect of abscisic acid, epibrassinolide, polyethylene glycol, naphthaleneacetic acid, 6-benzylaminopurine and arabinogalactan proteins. Euphytica 195, 369-382. Chupeau Y., Caboche M. and Henry Y. (1998). Androgenesis and haploid plants. (Berlin, Heidelberg.: Springer-Verlag). Kelliher T., Starr D., Wang W., McCuiston J., Zhong H., Nuccio M.L. and Martin B. (2016). Maternal Haploids Are Preferentially Induced by CENH3-tailswap Transgenic Complementation in Maize. Front Plant Sci 7. Kim M., Jang I.-C., Kim J.-A., Park E.-J., Yoon M. and Lee Y. (2008). Embryogenesis and plant regeneration of hot pepper (Capsicum annuum L.) through isolated microspore culture. Plant Cell Rep. 27, 425-434. Maluszynski M., Kasha K.J., Forster B.P. and Szarejko I. (2003). Doubled haploid production in crop plants. A manual. (Dordrecht, The Netherlands: Kluwer Academic Publishers). Palmer C.E. and Keller W.A. (2005). Overview of haploidy. En Haploids in crop improvement II, C.E. Palmer, W.A. Keller and K.J. Kasha, eds (Berlin Heidelberg: Springer-Verlag), pp. 3-9. Parra-Vega V., González-García B. and Seguí-Simarro J.M. (2013a). Morphological markers to correlate bud and anther development with microsporogenesis and microgametogenesis in pepper (Capsicum annuum L.). Acta Physiol. Plant. 35, 627-633. Parra-Vega V., Renau-Morata B., Sifres A. and Seguí-Simarro J.M. (2013b). Stress treatments and in vitro culture conditions influence microspore embryogenesis and growth of callus from anther walls of sweet pepper (Capsicum annuum L.). Plant Cell Tissue Organ Cult. 112, 353-360. Ravi M. and Chan S.W.L. (2010). Haploid plants produced by centromere-mediated genome elimination. Nature 464, 615-618. Rihova L. and Tupy J. (1999). Manipulation of division symmetry and developmental fate in cultures of potato microspores. Plant Cell Tissue Organ Cult. 59, 135-145. Rivas-Sendra A., Campos-Vega M., Calabuig-Serna A. and Seguí-Simarro J.M. (2017). Development and characterization of an eggplant (Solanum melongena) doubled haploid population and a doubled haploid line with high androgenic response. Euphytica 213, 89. Rivas-Sendra A., Corral-Martínez P., Camacho-Fernández C. and Seguí-Simarro J.M. (2015). Improved regeneration of eggplant doubled haploids from microspore-derived calli through organogenesis. Plant Cell Tissue Organ Cult. 122, 759-765. Rotino G.L. (1996). Haploidy in eggplant. En In vitro haploid production in higher plants, S.M. Jain, S.K. Sopory and R.E. Veilleux, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp. 115-141. Sanei M., Pickering R., Kumke K., Nasuda S. and Houben A. (2011). Loss of centromeric histone H3 (CENH3) from centromeres precedes uniparental chromosome elimination in interspecific barley hybrids. Proceedings of the National Academy of Sciences 108, E498-E505. Seguí-Simarro J.M., Bárány I., Suárez R., Fadón B., Testillano P.S. and Risueño M.C. (2006). Nuclear bodies domain changes with microspore reprogramming to embryogenesis. Eur. J. Histochem. 50, 35-44. Seguí-Simarro J.M. and Nuez F. (2005). Meiotic metaphase I to telophase II is the most responsive stage of microspore development for induction of androgenesis in tomato (Solanum lycopersicum). Acta Physiol. Plant. 27, 675-685. Seguí-Simarro J.M. and Nuez F. (2006). Androgenesis induction from tomato anther cultures: callus characterization. Acta Hort. 725, 855-861. Seguí-Simarro J.M. and Nuez F. (2007). Embryogenesis induction, callogenesis, and plant regeneration by in vitro culture of tomato isolated microspores and whole anthers. J. Exp. Bot. 58, 1119-1132. Seguí-Simarro J.M. and Nuez F. (2008a). How microspores transform into haploid embryos: changes associated with embryogenesis induction and microspore-derived embryogenesis. Physiol. Plant. 134, 1-12. Seguí-Simarro J.M. and Nuez F. (2008b). Pathways to doubled haploidy: chromosome doubling during androgenesis. Cytogenet. Genome Res. 120, 358-369. Shtereva L.A., Zagorska N.A., Dimitrov B.D., Kruleva M.M. and Oanh H.K. (1998). Induced androgenesis in tomato (Lycopersicon esculentum Mill). II. Factors affecting induction of androgenesis. Plant Cell Rep. 18, 312-317. Srivastava P. and Chaturvedi R. (2008). In vitro androgenesis in tree species: An update and prospect for further research. Biotechnol. Adv. 26, 482-491. Supena E.D.J., Muswita W., Suharsono S. and Custers J.B.M. (2006a). Evaluation of crucial factors for implementing shed-microspore culture of Indonesian hot pepper (Capsicum annuum L.) cultivars. Sci. Hort. 107, 226-232. Supena E.D.J., Suharsono S., Jacobsen E. and Custers J.B.M. (2006b). Successful development of a shed-microspore culture protocol for doubled haploid production in Indonesian hot pepper (Capsicum annuum L.). Plant Cell Rep. 25, 1-10. Touraev A., Pfosser M. and Heberle-Bors E. (2001). The microspore: A haploid multipurpose cell. Adv. Bot. Res. 35, 53-109. Zagorska N.A., Shtereva A., Dimitrov B.D. and Kruleva M.M. (1998). Induced androgenesis in tomato (Lycopersicon esculentum Mill.) - I. Influence of genotype on androgenetic ability. Plant Cell Rep. 17, 968-973. |




|
© 2019 Cell Biology Group. |
Legal Notice |